Your new post is loading...
Your new post is loading...

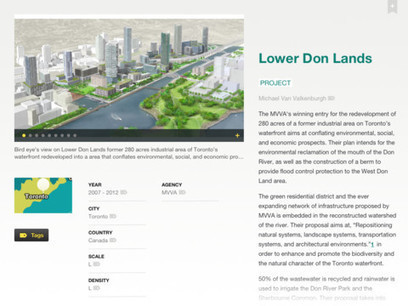
Abstract Artificial light at night (ALAN) is an increasingly important driver of global change. Lighting directly affects plants, but few studies have investigated indirect effects mediated by interacting organisms. Nocturnal Lepidoptera are globally important pollinators, and pollen transport by moths is disrupted by lighting. Many street lighting systems are being replaced with novel, energy‐efficient lighting, with unknown ecological consequences. Using the wildflower Silene latifolia, we compared pollination success and quality at experimentally lit and unlit plots, testing two major changes to street lighting technology: in lamp type, from high‐pressure sodium lamps to light‐emitting diodes, and in lighting regime, from full‐night (FN) to part‐night (PN) lighting. We predicted that lighting would reduce pollination. S. latifolia was pollinated both diurnally and nocturnally. Contrary to our predictions, flowers under FN lighting had higher pollination success than flowers under either PN lighting or unlit controls, which did not significantly differ from each other. Lamp type, lighting regime, and distance from the light all significantly affected aspects of pollination quality. These results confirm that street lighting could affect plant reproduction through indirect effects mediated by nocturnal insects, and further highlight the possibility for novel lighting technologies to mitigate the effects of ALAN on ecosystems. Introduction Artificial light at night (ALAN) represents one of the fastest growing anthropogenic environmental changes globally, with background sky brightness increasing by 6%/yr (Hölker et al. 2010). Although often associated with urbanization, artificial light also affects natural and semi‐natural ecosystems, with temperate ecosystems experiencing some of the fastest increases in exposure (Bennie et al. 2015). The nature of artificial lighting is also changing rapidly. An increasing drive for energy efficiency, cost reduction, and improved visibility has led to the widespread implementation of novel lighting technologies for street lighting. Most notably, part‐night (PN) lighting regimes are being introduced; whereby, lights may be switched off or dimmed at certain times of night (Stone et al. 2015). Also, the incumbent lamp types, especially high‐pressure sodium (HPS) lights, are being replaced by arrays of light‐emitting diodes (LEDs; Gaston et al. 2015). The uptake of LED lighting can alter assemblages of nocturnal invertebrates (Davies et al. 2017) and is recognized as a key emerging threat to biodiversity in urban ecosystems due to the greater emission of blue light by LEDs compared to incumbent lighting technologies (Stanley et al. 2015). Artificial light has the potential to affect every level of biological organization from cells to communities (Gaston et al. 2015), and may affect diurnal communities as well as nocturnal (Manfrin et al. 2017). Despite this, little attention has been paid to the potential disruptive effect that artificial light may have on ecosystem services provided by nocturnal organisms (Lewanzik and Voigt 2014). In a recent review, Bennie et al. (2016) highlighted the need to investigate the indirect effects of light on plants, mediated by herbivores, pollinators, and other interacting organisms. A growing body of evidence suggests that nocturnal moths play a globally important role as pollinators (Banza et al. 2015, Macgregor et al. 2015). Individually, moths are known to be affected by artificial light through a wide range of mechanisms (Macgregor et al. 2015, van Geffen et al. 2015a, b, Wakefield et al. 2015). Their interactions with flowers can be directly affected through reduced feeding (van Langevelde et al. 2017), and we have previously found that pollen transport by nocturnal moths can be disrupted by the presence of full‐night (FN) lighting by HPS street lights (Macgregor et al. 2017): Moths at lit sites were attracted upwards to street lights, reducing the time they spent at the level of the field margin, where interactions with flowers might take place, compared to unlit sites. We found that moths at lit sites were significantly less likely to be transporting pollen on the proboscis, and there was some evidence this might lead to significant overall reductions in the quantity and diversity of pollen transported nocturnally by moths (Macgregor et al. 2017). Indeed, recent work has shown that fruit set is reduced in Cirsium oleraceum (L.) Scop. under FN LED street lighting (Knop et al. 2017). However, reduced pollen transport may not necessarily lead to reduced pollination (Ballantyne et al. 2015) for all plant species or under all types of street lighting. In this study, we assessed the effects of artificial lighting, including the relative effects of two changes in lighting technology (from HPS to LED, and from FN to PN lighting), upon pollination by nocturnal pollinators, using the plant white campion Silene latifolia Poir. as a bioassay. Silene latifolia is a widespread archaeophytic wildflower in the United Kingdom (Baker 1947); it often grows on road verges and therefore is prone to exposure to artificial light from street lights (Bennie et al. 2016). Pollination is conducted by both diurnal and nocturnal insects, and pollen is transported nocturnally by a broad range of generalist moth species of several families (Macgregor et al. 2017). As a result, the species has been used as a model organism in studies of nocturnal pollination by moths in Europe and North America (Barthelmess et al. 2006, Hahn et al. 2015). Moths are significantly better both at pollinating female flowers and at dispersing pollen over distance than diurnal visitors (Young 2002, Barthelmess et al. 2006), but previous studies have not investigated whether the two groups act to provide complementarity or redundancy to the pollination system of S. latifolia. Pollination is a critical ecological process as well as a valuable ecosystem service (Kearns et al. 1998), and understanding the contributions of all guilds of pollinators (including those that are nocturnal) is of key importance. We addressed three main questions in this study. Firstly, we investigated the relative contributions of diurnal and nocturnal pollinators to the pollination of S. latifolia, aiming to establish whether the two pollinator guilds show redundancy. Secondly, we investigated the effect of FN and PN street lighting on pollination of S. latifolia, compared to unlit controls. Based on our previous finding that pollen transport is significantly reduced in the presence of HPS street lighting (Macgregor et al. 2017), we predicted that both pollination success and quality (the proportion of flowers setting seed, and the number/mass of seeds per flower, respectively) would be reduced in the presence of FN lighting, due to a reduction in pollen transport by nocturnal moths. Our expectation was that the hypothesized reduction in pollination under FN lighting would be smaller under PN lighting, compared to unlit controls, because the disruption to pollen transport caused by lighting would not be imposed for the latter part of the night. Thirdly, we investigated the effects of different types of street lighting technology on pollination of S. latifolia, comparing FN lighting to PN lighting and HPS lights to LEDs using a fully crossed experimental design. As discussed above, we predicted that PN lighting would have a lesser impact on pollination, and therefore greater pollination success and quality than FN lighting. Previous studies have variously reported LED lighting to be both more (Pawson and Bader 2014) and less (Huemer et al. 2010) attractive to nocturnal invertebrates than, or to have no effect relative to (Wakefield et al. 2018), HPS; therefore, while we tested for an effect of change in lighting type, we did not make a prediction for the direction of any such effect. Materials and Methods Study organisms We conducted a bioassay of pollination using Silene latifolia Poir. (=S. alba (Mill.) E.H.L. Krause; Caryophyllaceae), a dioecious wildflower native to Eurasia, archaeophytic in the UK (Baker 1947), and introduced in North America and Australasia (McNeill 1977). S. latifolia can receive pollination from both diurnal and nocturnal pollinators (Young 2002), but it is not established whether the two guilds of pollinators are redundant or complementary. S. latifolia plants were grown from seed in a glasshouse at Thwaite Botanical Gardens (University of Hull), Cottingham, East Riding of Yorkshire, UK. Seeds were germinated and individual plants pricked out into 7 × 7 cm pots. Upon emergence of flowers, male and female plants were identified. Flowering shoots of female plants were supported with 120‐cm bamboo canes, and flowers were individually bagged using 7 × 9.4 cm organza bags to prevent access by pollinators prior to the start of the experiment. Male plants were supported with 60‐cm bamboo canes and flowers left open. Female plants were selected for each replicate of the experiment (henceforth, experimental run) according to the availability of open, virgin flowers, such that every plant had at least one flower that was exposed to pollinators for the full four days and nights of each experimental run; however in some cases, additional flowers may have opened during the experimental run, which would have been exposed for a subset of this time. All flowers that were fully mature at the end of each experimental run were included in the final dataset. As flowers of S. latifolia are open from late afternoon until mid‐morning (Young 2002), any fully mature flower had to have first opened on the previous evening or earlier, and had therefore been exposed to one full cycle of pollination (including nocturnal and diurnal pollinators) by that point. As female plants continuously produced flowers throughout the experimental period, some individual plants were re‐used across multiple experimental runs (being randomly re‐assigned to a treatment, which therefore may have been either the same as on previous uses or different), depending on the availability of previously unused female plants in flower at the commencement of each experimental run. In total, 169 individual female plants were used across the four experimental runs; 111 plants were used in one run only, 46 plants were used twice, 11 plants were used three times, and one plant was used in all four runs. Field site The experiment was conducted in field margins at Molescroft Grange Farm, a 600‐ha arable farm managed sustainably under the Linking Environment and Farming (LEAF) scheme, near Beverley, East Riding of Yorkshire, UK (53°51′44″ N 0°25′14″ W). Field margins, consisting of strips of non‐crop semi‐natural habitat between the crop and the boundary features, are a key feature of agroecosystems in the UK and provide refuges for many species of plants and animals (Marshall and Moonen 2002). The field margins at Molescroft Grange Farm were ~3 m wide and bounded on one side by the crop and on the other by a 2 m deep ditch with a hedge behind. Margins were floristically dominated by grass, but also contained a range of wildflower species. Although no naturally occurring S. latifolia was observed on site, the species is present in the region. However, S. dioica (L.) Clairv. was abundant at the study site; these two species are known to hybridize (Minder et al. 2007). Five experimental plots were established in the margins of two adjacent fields (Appendix S1: Fig. S1), one containing wheat and the other a grass mix for hay production. Plots were >300 m apart from each other and separated where possible by visual barriers such as trees or hedges; as this distance was substantially greater than the maximum reported distance from which moths may be attracted to lights (Baker and Sadovy 1978, Beck and Linsenmair 2006, Truxa and Fiedler 2012, van Grunsven et al. 2014, Merckx and Slade 2014), we considered the effects of the lighting treatment at each plot to be independent of treatments at other plots. Experimental runs Each experimental run lasted for four days and nights, starting and ending at roughly midday; we aimed to allow sufficient time for some flowers to be pollinated in each treatment, but also to keep the experiment sufficiently short that not all flowers would be pollinated, allowing us to investigate any variation in pollination success between treatments. Experimental runs were initiated when medium‐range weather forecasts indicated that conditions would be favorable for pollinator activity (i.e., conditions associated with high barometric pressure) for the majority of the run. The actual weather conditions experienced during the experimental run often differed from those forecasts, but were broadly consistent across all experimental runs (Appendix S1: Table S2). The experiment was repeated four times in total (beginning on 26 June, 15 July, 30 July, and 14 August 2015, respectively), with each treatment being moved to a different plot for each repeat, such that over the course of the experiment, each lighting treatment (including the unlit control) was tested at four out of the five experimental plots. Caging treatments During each experimental run, we placed one group of four female plants each at 0, 10, and 20 m in a straight line along the field margin from the center of each plot, where lighting treatments were placed; therefore, each experimental run used 60 female plants (12 per lighting treatment). Within each group, the four female plants were arranged in a square, with pots separated by 7 cm, and four male plants (to act as pollen donors) in a larger square surrounding the females at ~50 cm distance (sufficient to completely exclude the possibility of pollination by direct contact between male and female flowers). To manipulate access to flowers by insect visitors, female plants were completely enclosed to ground level in net cages. Cage frames were constructed using 1.8‐m bamboo canes with stripwood spacers to give a footprint of 25 × 15 cm, such that two adjacent plants could be simultaneously enclosed in a single cage. Insects were excluded from cages using 0.6‐mm polyethylene mesh (Gardman InsectGard, Gardman, Peterborough, UK), secured to the frame using cable ties; mesh was wrapped 1.5 times around the cage to ensure full overlap at the joining edge. One female plant from each group of four was subjected to each of the four caging treatments: always caged (a control treatment; henceforth fully caged), always uncaged (openly pollinated), uncaged during the day only (diurnally pollinated), and uncaged during the night only (nocturnally pollinated). Plants in the diurnally pollinated treatment were caged from dusk until dawn, and plants in the nocturnally pollinated treatment were caged from dawn until dusk for the duration of each experimental run. Groups of plants were enclosed in a 1 m high chicken‐wire cylinder of ~45 cm diameter, which protected plants from grazing by rabbits or deer and offered some support to plants and cages under windy conditions. As most flowers were roughly the same height as the top of the chicken‐wire cylinder, we expected the majority of insects to access flowers over the top of the cylinder, unhindered by the wire; additionally, due to the large diameter of holes in the wire, it is likely that smaller insects would have been able to fly through the holes. Lighting treatments In each experimental run, we applied one unlit control treatment and four different lighting treatments across the five plots. We tested the two current major transitions in UK street lighting technology: the change from high‐pressure sodium lights to light‐emitting diodes (LEDs), and the change from FN lighting to PN lighting (where lights are switched off at midnight); therefore, the four lit treatments were HPS‐FN, HPS‐PN, LED‐FN, and LED‐PN lighting. We constructed 4 m high lighting rigs in the field margins (4 m is the minimum height for highway lighting in the UK and a common height for street lighting on minor roads in rural settings). Floodlights were mounted on tripod VHF‐radio masts (Sandpiper Aerials, Aberdare, UK). We used commercially available floodlights, purchased from SCL Direct (Sheffield, UK); for HPS lights, we used a 70‐W HPS‐bulb floodlight with 5300 lm luminous flux (output); and for LED lights, we used a 50‐W LED‐array floodlight with 5000 lm output. These lights were selected to have similar output to each other and are typical of lighting used at this height in rural settings (e.g., see British Standards Institution 2013, Hampshire County Council 2016), but differed substantially in their spectral composition (Fig. 1), measured using a fully corrected spectrometer with neutral density filters (Jobin Yvon Fluorolog tau‐4, Jobin Yvon, Edison, New Jersey, USA). The lights also differed slightly in their light intensity at ground level (Fig. 1; measured using a handheld light meter: Holdpeak HP‐881C, Holdpeak, Hong Kong): Light intensity was slightly but significantly higher underneath the LED lights (F = 9.38, P = 0.018) and decreased with increasing distance from the lights (F = 1250.6, P < 0.001). Although light intensity was higher at 15–20 m distance from the HPS lights than at the same distance from the LED lights, indicating greater light scatter from the HPS lights (Appendix S1: Table S1), there was no significant interaction between lamp type and distance (F = 0.02, P = 0.895). Lights were turned on and off automatically by a combination of two switches. All lights were turned on at dusk by a photocell switch; this switch also turned off the lights in the FN treatments at dawn. Lights in the PN treatments were turned off at midnight by a timer switch. Each light was powered using a 1500‐W petrol generator (Rhyas RH1500; Rhyas, Leominster, UK). Assessment of pollination At the finish of each experimental run, female plants were returned to the greenhouse and all open, fully mature flowers were again enclosed in bags to prevent further access by any insects that were potentially present in the greenhouse. Flowers were left for one week to develop into seed capsules, were removed, and then stored individually. Seed capsules were oven‐dried for 48 h at 75–80°C. Pollination success was assessed by the development of one or more seeds in the seed capsule and recorded as a binary variable (success = 1, failure = 0). Further measurements of total number of seeds per seed capsule (seed count) and total mass of the seed contents of each seed capsule (seed mass, g) were recorded for all pollinated, unparasitized (see following paragraph) flowers. These measures have been used in previous studies as indicators of pollination quality in S. latifolia and can differ between diurnal and nocturnal pollinator guilds, possibly due to the greater age of stigmas when exposed to diurnal pollination, after initial anthesis at or around dusk (Young 2002). We recorded seed mass of the entire seed capsule, rather than the average mass of individual seeds because individual seeds were very small and, although some variation in size was apparent, we did not consider that the average size of individual seeds was a true measure of pollination quality of the whole floral unit. In some cases, flowers were found to have been parasitized by Lepidopteran larvae feeding internally on developing seed capsules (46 out of 194 successfully pollinated flowers). Seed count and seed mass could not be recorded for these flowers. Larvae were observed in the glasshouse to occasionally move between flowers and to be able to move in and out of the closure of the organza bags along the flower stem. Seven (15.2%) of the parasitized flowers had been exposed only to diurnal pollinators in the field, and two (4.3%) were fully caged and had not been exposed to either diurnal or nocturnal pollinators in the field. The frequency of parasitism was not significantly related to caging treatment (Pearson's chi‐square test, χ2 = 3.77, df = 3, P = 0.288). Therefore, it could not be determined beyond reasonable doubt whether each larva had developed on the parasitized flower from eggs laid in the field or had moved onto that flower from another, after it was returned to the glasshouse. For that reason, parasitization was not itself recorded as a variable, but it was assumed that all parasitized capsules had been successfully pollinated, as non‐pollinated flowers would not contain seeds for larvae to feed on. Statistical analysis All statistical analyses were conducted in R version 3.4.4 (R Core Team 2018), and figures were plotted using the package ggplot2 (Wickham 2009). R code used in the analysis is archived at dx.doi.org/10.5281/zenodo.1743917. We analyzed three dependent variables: the number of pollinated and non‐pollinated flowers per plant (pollination success; effectively the probability of each individual flower within a given plant being successfully pollinated), seed count per pollinated seed capsule, and total seed mass per pollinated seed capsule. Generalized linear mixed‐effects models (GLMMs) were constructed to test for differences in pollination success and quality between flowers under different caging and lighting treatments, using the package lme4 (Bates et al. 2014). Models to analyze pollination success were constructed using a binomial error distribution, and pollination quality using a negative binomial error distribution (to correct for overdispersion). The experimental run (1–4) was included as a categorical random effect in all models. In addition to the main fixed effects (lighting treatment, caging treatment, and distance from the light), we included a numerical covariate, number of uses (i.e., whether the data were recorded from the first, second, third, or fourth use of each plant), in all models to account for the possibility that either (1) older plants might be less productive or (2) plants that had previously invested in setting seed might be less productive. We also separately analyzed the relationship between number of uses and each dependent variable, using GLMMs with both the individual plant ID and the experimental run included as random effects. Significance of fixed effects in all GLMMs was tested using likelihood ratio tests. The analysis addressed three distinct questions in order to fully explore the relative effects of changing lighting regime (from FN to PN, including a comparison to unlit controls) and lamp type (from HPS to LED), because our five‐level categorical lighting treatment could not be full‐factorial (i.e., it was impossible to have a treatment that was simultaneously FN or PN for light regime, but unlit for lamp type, or vice versa), and therefore might mask subtle effects such as interactions between distance from the light and either lamp type or lighting regime. Specifically, we analyzed: (1) the overall effects of the five‐level lighting treatment on, and relative contributions of diurnal and nocturnal pollinators to, the pollination of S. latifolia; (2) the effect of FN street lighting (HPS‐FN and LED‐FN treatments combined) and, separately, PN street lighting (HPS‐PN and LED‐PN treatments combined) on pollination of S. latifolia, compared to unlit controls; and (3) the effects of different types of street lighting technology (HPS vs. LED, and FN vs. PN) on pollination of S. latifolia, compared to each other in a four‐way full‐factorial model (for which it was necessary to exclude data from unlit controls). For each question, we analyzed effects on all three dependent variables. To assess the overall effects of the five‐level lighting treatment on, and the relative contributions of diurnal and nocturnal pollinators to, the pollination of S. latifolia (question 1), we constructed a full model to describe all biologically plausible interaction terms between variables, with the fixed‐effects structure: pollination ~ (lighting treatment × caging treatment) + (lighting treatment × distance) + number of uses. Lighting treatment was represented by a five‐level categorical variable. Where interaction terms were not found to be significant, models were reconstructed with interactions removed and re‐tested. To test for differences in pollination success and quality between the different caging treatments, we conducted post‐hoc Tukey HSD tests on the final model, using the package multcomp (Hothorn et al. 2008). Additionally, we carried out power analyses on the models constructed for question (1), to assess their ability to detect significant interactions (of the observed effect size) between lighting treatment, caging treatment, and distance from the light (Appendix S1: Table S3). For each of our three dependent variables (pollination success, seed count, and seed mass), we conducted four power analyses, with the following fixed‐effects structures: (1) pollination ~ lighting treatment + caging treatment + distance + number of uses; (2) pollination ~ (lighting treatment × caging treatment) + (lighting treatment × distance) + number of uses; (3) pollination ~ (lighting treatment × caging treatment) + distance + number of uses; and (4) pollination ~ (lighting treatment × distance) + caging treatment + number of uses. Power of each model was the percentage of 1000 simulated datasets for which a significant effect of lighting (or its relevant higher‐order interaction term) was found. Data were simulated using the sim.glmm function from the R package GLMMmisc (Johnson 2016), using parameters generated by fitting the model structure in question to the observed data. Where lighting treatment was used in more than one interaction term in the same model, we estimated power of both interaction terms using the same set of 1000 simulations. The results of the power analyses are given in Appendix S1: Table S3. To assess the effects of FN lighting and PN lighting, compared to unlit controls, on the pollination of S. latifolia (question 2), we separately constructed models using two subsets of data, to test the effect of FN light compared to unlit controls with data from PN lit plants excluded, and vice versa. Models were constructed with the fixed‐effects structure: pollination ~ (lighting treatment × caging treatment) + (lighting treatment × distance) + number of uses; as before, non‐significant interaction terms were removed and the models re‐tested in their absence. Finally, to assess the effects of different types of lighting treatment (HPS vs. LED and FN vs. PN) on pollination of S. latifolia (question 3), we constructed models using a third subset of data, with unlit controls excluded. Models were constructed with the fixed‐effects structure: pollination ~ (light regime × caging treatment) + (lamp type × caging treatment) + (light regime × lamp type) + (light regime × distance) + (lamp type × distance) + number of uses, and non‐significant interaction terms removed as before. Results Effects of caging and lighting treatments on pollination Across all experimental runs and plants, we sampled 417 flowers of S. latifolia from 169 individual plants. A total of 194 flowers were successfully pollinated and of these 46 flowers (from 39 different plants) were parasitized, allowing us to measure seed count and weight for 148 flowers. The mean seed count was 74.16 (range: 4–354), and the mean seed weight was 29.0 mg (range: 3.9–128.0 mg). Flowers of S. latifolia were successfully pollinated by both diurnal and nocturnal pollinators (Fig. 2). The highest pollination success was among flowers that were openly pollinated (always uncaged) and had access to both diurnal and nocturnal pollinators (72.0% of such flowers were pollinated). Comparing diurnally pollinated flowers (uncaged only during the day) and nocturnally pollinated flowers (uncaged only during the night), nocturnally pollinated flowers had higher rates of pollination (52.8%, compared to 37.6% of diurnally pollinated flowers). Therefore, combining these probabilities, the expected rate of pollination for flowers open to both diurnal and nocturnal pollinators was (1 − (1 − 0.528) × (1 − 0.376)) = 70.5%, which was very close to the observed value of 72%. This indicates that, in our study system over a four‐day period of exposure to pollinators, diurnal and nocturnal pollinators of S. latifolia were complementary (Blüthgen and Klein 2011) but did not provide redundancy. Two fully caged flowers set seed; a previous study of S. latifolia also recorded low levels of seed set in flowers that had been fully caged and suggested that this might be attributable to small insects (e.g., thrips) that are able to pass through the mesh of the net cages (Young 2002). Because the rate at which pollination occurred in the fully caged treatments was non‐zero, we retained these data in subsequent statistical analyses; nevertheless, we are confident that this rate was sufficiently low that our results are robust. We found that both caging treatment (Fig. 2) and the five‐level categorical lighting treatment (Fig. 3) significantly affected pollination success of S. latifolia (Appendix S1: Table S4), although we found no significant interaction between the two. Surprisingly, we also found no significant effect of distance from the light (0–20 m) or of the interaction between lighting treatment and distance on pollination success. In the full model, there was no significant effect of lighting treatment, caging treatment, distance, or any interaction between the three on either seed count or seed mass. We found that many of the differences in pollination success between the four caging treatments were significant (Appendix S1: Table S5). Specifically, the pollination success of openly pollinated flowers was significantly higher than all other treatments, and both diurnally pollinated and nocturnally pollinated flowers had significantly higher pollination success than fully caged flowers; however, there was no significant difference between pollination success in diurnally pollinated and nocturnally pollinated flowers (Fig. 2). Effects of lighting, compared to unlit controls To fully understand the effects of lighting on pollination success and quality in S. latifolia, we also analyzed the effects of specific lamp types and lighting regimes relative to each other and to unlit controls, because our five‐level categorical lighting treatment could not be full‐factorial. Comparing FN lighting to unlit controls, pollination success was significantly higher under FN lighting (Fig. 4). There was no significant difference in seed count or seed mass between flowers under FN lighting and flowers in unlit controls (Appendix S1: Table S6). Comparing PN lighting to unlit controls, there was no significant difference in pollination success, seed count, or seed mass between flowers under PN lighting and flowers in unlit controls (Fig. 4). Effects of different types of street lighting technology Comparing between lighting regimes (FN vs. PN) and lamp types (HPS vs. LED), we found that pollination success was significantly higher under FN lighting than under PN lighting (Fig. 5). Lamp type and lighting regime had a significant, interacting effect on seed count (Fig. 5); specifically, seed count did not differ between HPS lights and LEDs when under FN lighting, but under PN lighting, seed count was higher under HPS lights and lower under LEDs. Lamp type and distance from the light had a significant, interacting effect on seed mass (Fig. 6); specifically, seed mass increased with increasing distance from HPS lights, but decreased with increasing distance from LED lights (Appendix S1: Table S7). Effect of re‐using individual plants The number of uses of each plant significantly affected seed count and dry mass of seeds per seed capsule (Appendix S1: Fig. S2), and number of uses was also found to significantly affect seed count and seed mass as a covariate in most models (Appendix S1: Tables S3, S5–6); seed count and seed mass reduced with each subsequent use of a plant. However, although there appeared to be a similar negative trend in pollination success as the number of uses increased (Appendix S1: Fig. S2), this was not significant either when analyzed independently (Appendix S1: Table S8) or as a covariate in any model. Discussion Overview Although evidence exists for a reduction in pollen transport and pollination by nocturnal moths at lit sites, compared to unlit controls (Knop et al. 2017, Macgregor et al. 2017), this study provides the first direct evidence that changes in street lighting technology can alter the effect of ALAN on pollination by nocturnal invertebrates. We found that S. latifolia plants in the vicinity of FN lighting appeared to have enhanced pollination success compared to plants in unlit controls; this result was in direct contrast to the reduction in pollination success and quality under lighting that we had hypothesized. We also found that PN lighting did not appear to have any strong effect on pollination success or quality, and therefore may have less biological impact than FN lighting, as predicted. Although we did not find any difference in pollination success between flowers under HPS lights and those LEDs, we found lamp type and lighting regime had an interacting effect on seed count, while lamp type and distance from the light had an interacting effect on seed mass. Relative contributions of diurnal and nocturnal pollinators Our experiment showed that both diurnal and nocturnal pollinators contribute to the pollination of S. latifolia, supporting the findings of previous studies (Young 2002). Nocturnally pollinated flowers appeared to have higher pollination success than diurnally pollinated flowers, though the difference was not significant. However, both treatments had significantly lower pollination success than openly pollinated flowers. This indicates that nocturnal and diurnal pollinators were complementary to each other in the pollination of S. latifolia (Blüthgen and Klein 2011). There was no redundancy between the guilds and so pollination success in this species is greatest in the presence of both guilds. Had flowers been allowed to remain in the field for a longer duration until they had fully completed flowering, or had the experiment been conducted at a site with a greater abundance and diversity of pollinators, it is possible that a higher rate of pollination may have led to the emergence of a degree of redundancy. Nevertheless, multiple paternity, which may increase female and offspring fitness by allowing pollen competition, has been demonstrated in S. latifolia (Teixeira and Bernasconi 2007). Even if flowers were not pollen‐limited (i.e., were successfully pollinated by at least one pollinator in all cases), visits from additional pollinating insects might still increase fitness by increasing the number of pollen donors from which pollen is received. Therefore, our findings suggest that a reduction in either guild of pollinators would impact upon pollination, and hence plant reproduction, given that S. latifolia is a biennial or short‐lived perennial (Baker 1947). Rates of pollination success in the fully caged control treatment were consistently very low across all replicates (Appendix S1: Table S5), indicating that our caging was mostly effective at excluding insect pollinators. Nevertheless, there were still low levels of pollination recorded in caged flowers. It is therefore possible that some diurnal visitors pollinated flowers in the nocturnally pollinated treatment, and vice versa; this might have introduced some bias to the data. Similarly, our inability to prevent seed‐feeding moth larvae from accessing the seed capsules or moving between flowers after plants had been returned to the glasshouse excluded the possibility of recording seed parasitism as an experimental variable. Several examples have been documented of pollinating seed parasitism (or nursery pollination) interactions involving S. latifolia and closely related plant species (Kephart et al. 2006), whereby adult moths (including males; see Labouche and Bernasconi 2010) pollinate the flowers of S. latifolia but females also oviposit upon them, and larvae feed internally on the developing seedheads. Therefore, we initially presumed these larvae to be either Hadena bicruris (Hufnagel, 1766) or Sideridis rivularis (Fabricius, 1775; =H. rivularis), both of which have well‐known nursery pollination interactions with S. latifolia (Kephart et al. 2006). However, several larvae were collected while apparently feeding on seed capsules in the glasshouse; of these, one was successfully reared to adulthood and the imago identified as Lacanobia oleracea (Linnaeus, 1758). This species has not previously been recorded to feed on S. latifolia seed capsules (Crafer 2005) but is a noted glasshouse pest of tomatoes (Gatehouse et al. 1997), including internal feeding on fruits (Jarrett and Burges 1982), and so may plausibly be responsible for the observed damage to our seed capsules. Adults of L. oleracea were light‐trapped at the field site (although light traps were not operated during the same nights when experimental runs were taking place), so it is possible that larvae had developed on the parasitized flowers from eggs laid in the field. It would therefore be of interest to analyze whether the frequency of pollinating seed parasitism is affected by artificial light, not least because this variable can be directly linked to interactions between nocturnal moths and flowers. Effects of lighting on pollination We found that artificial light significantly affected the pollination success of S. latifolia (Fig. 3). However, we did not find any significant interaction between lighting treatment and either caging treatment or distance from the light in our full models, despite the fact that both interactions are biologically plausible (though we did find that lamp type and distance from the light had a significant, interacting effect on seed mass; Fig. 6). However, power analyses suggested that we had relatively low power to detect such interaction terms for both pollination success and seed count (roughly 10–20% lower than for the equivalent models with interaction terms not included, and in most cases also lower than the ideal of >80% power; Appendix S1: Table S3), so it is possible that these interactions do exist in truth, and might have been detected with a larger dataset. Alternatively, it is possible that the effect of distance from the light operates at a larger scale than we tested in this study (0–20 m); certainly, most species of larger moths are capable of flying many times more than 20 m in the course of a single night (Jones et al. 2016). Further work is necessary to resolve these points. Pollination success increased under FN lighting compared to unlit controls, contrary to our hypothesis that pollination would be reduced under artificial light. However, there was no difference in pollination success under PN lighting compared to unlit controls, which appears to support our hypothesis that PN lighting would cause less ecological disruption (relative to unlit controls) than FN lighting. The cause of these observed patterns is not clear. Potentially, lights could act as ecological traps, retaining an artificially high density of moths in their vicinity and leading to locally elevated rates of flower visitation (one of the possible effects of lighting proposed by Macgregor et al. 2015). However, this would appear to be contradicted by the findings of our previous study, which indicated a reduction in overall pollen transport at lit sites (Macgregor et al. 2017), and of Knop et al. (2017), which found a reduction in fruit set of Cirsium oleraceum (Asteraceae) under FN LED lighting. This conflict raises the possibility that ALAN could create imbalances among plant communities, with some species benefitting and others suffering from exposure to street lights. One mechanism by which pollination of S. latifolia could increase, without the same increase being replicated for other species, is that lighting might increase the visibility of S. latifolia flowers to moths. S. latifolia is a relatively conspicuous white flower, and our plants were generally taller than the surrounding sward in the experimental plots; lower‐growing plants or those of different colors might not experience the same benefit of proximity to a light. Under PN lighting, plants would still be exposed to a period of natural darkness, during which any benefits of exposure to light might not continue. Aspects of artificial lighting also affected both the number of seeds produced per seed capsule and the total dry mass of seeds per seed capsule, though the two would be expected to correlate. Specifically, an interaction between lamp type and lighting regime meant that seed count increased under PN HPS lighting and decreased under PN LED lighting, relative to unlit controls (Fig. 3) and both FN lighting treatments. However, an interaction between lamp type and distance meant that seed mass increased with distance from the light under HPS lighting but decreased with distance from the light under LEDs (Fig. 6). Again, the cause of these effects is unclear, and it should be noted that both effects were non‐significant when lighting was treated as a five‐level categorical variable (Appendix S1: Table S4). Possibly, these effects could be the result of changes in duration of feeding by moths under different lamp types; moths spend less time feeding under artificial light than in darkness, and the effect is strongest for lighting containing a high proportion of short wavelengths (van Langevelde et al. 2017), such as many commercially available LEDs, including those used in our study (Fig. 1). The strength of any effect of light can clearly be expected to reduce with distance from the light. However, it is not clear how such effects would interact with lighting regime. A physiological response by the plant (i.e., increased photosynthesis under lighting treatments) appears unlikely because seed capsules mostly developed after all flowers were returned to the glasshouse, when they were no longer exposed to the experimental lights. There is a possibility that differences between lit treatments and unlit controls could have been influenced by low‐level noise and/or air pollution from the petrol generators used to power the experimental lights, as generators were not operated at the unlit control plots. Nevertheless, further research to understand the factors that drive this observed interaction between lamp type and lighting regime is important. Regardless of the cause of the observed interactions, our finding that the direction of the effects of HPS lights and LEDs can interact with other factors (here, light regime and distance from the light) is interesting in the context of several recent studies into the relative attractiveness of LEDs and other lamp types to nocturnal invertebrates (Huemer et al. 2010, Pawson and Bader 2014, Wakefield et al. 2018). Although these studies all compared the abundance of insects attracted to LEDs and other lamp types, including HPS, they together report that LED does not differ in attractiveness from HPS (Wakefield et al. 2018), or that LED is more attractive than HPS (Pawson and Bader 2014), or that LED is less attractive than HPS (Huemer et al. 2010). Our results may partially explain this conflict, by suggesting instead that the relative ecological impact of the two lighting types can be substantially altered by other, related variables, such as the duration of lighting. For this study, we used standard, commercially available LED floodlights, with typical spectral composition (Fig. 1); unlike other major lighting types, the spectral composition of LEDs can be customized, and previous work has demonstrated that this can reduce their attraction to insects (Longcore et al. 2015), potentially mitigating their ecological impact. The increase in energy efficiency from use of LEDs may encourage the uptake of this technology (Gaston et al. 2014), possibly at the cost of increased global levels of light pollution (Kyba et al. 2017), although carefully designed LED lighting systems can instead reduce light pollution (Kyba et al. 2018). Conclusions Viewed as a whole, the effects we have observed suggest an increase in nocturnal visitation of S. latifolia under FN lighting (compared to unlit controls) that was not detected under PN lighting. An interaction between lamp type (HPS vs. LED) and lighting regime affected the number of seeds produced per seedhead, while the interaction between lamp type and distance from the lamp affected the total dry mass of those seeds. We found no further significant effect of distance from the light in any other analysis within this study, although testing the effect of lighting over a greater range of distances is an important direction for future study and might clarify to what distance the effect of light can permeate. These findings lend further support to previous studies suggesting that PN lighting regimes may provide the best solution to reducing the influence of artificial night lighting on nocturnal wildlife (Azam et al. 2015) and mitigating its ecological impacts. While our study, based on a single plant species, appears to suggest that plants may benefit from being situated under FN lighting, comparing this unexpected result to Knop et al. (2017) demonstrates the potential for ALAN to disrupt pollination systems across the community of plants, disproportionately strengthening some interactions and weakening others (Macgregor et al. 2015). Community‐level studies would be valuable to identify potential winners and losers from lighting, to determine the exact mechanisms underpinning the effects we have identified, and to understand how best to mitigate negative impacts. Acknowledgments This work was supported by the Natural Environment Research Council and Butterfly Conservation (Industrial CASE studentship awarded to C.J.M., Project Reference: NE/K007394/1). We thank V. Swetez for growing the plants and the workshop staff at the University of Hull for constructing the lighting rigs. We are grateful to T. Hall for kindly allowing us to conduct the experiment at Molescroft Grange Farm, Beverley. We thank S. Amos, S. Bavin, and V. Scott for assistance in the laboratory and glasshouse. We thank A. Harriman of the Molecular Photonics Laboratory, Newcastle University, for measuring the spectral output of our experimental lights, and M. Brett of Cockle Park Farm, Newcastle University, for assisting with measuring light intensity under the experimental lighting rigs. All authors contributed to designing the experiment, analyzing the data, and preparing the manuscript. Field and laboratory work was conducted by C.J.M. Supporting Information Literature Cited

|
Scooped by LIGHTING |

No comment yet.
Sign up to comment